
Avances en Psicología Latinoamericana
ISSN:1794-4724 | eISSN:2145-4515
ISSN:1794-4724 | eISSN:2145-4515

Animal Model of Cognitive Decline and Neural Transplantation of Stem Cells as a Prospective Therapy
Modelos animales de degeneración cognitiva y uso de transplantes neurales de células madre como terapia
Modelos animais de declínio cognitivo e uso de transplantes neurais de células-mãe como terapia
Leticia Domingues Bertuzzi, Patrícia Santos da Silva, Élida Fluck Pereira Neto, Rafael Rafael Nicolaidis, Alcyr Alves de Oliveira Jr
Animal Model of Cognitive Decline and Neural Transplantation of Stem Cells as a Prospective Therapy
Avances en Psicología Latinoamericana, vol. 35, no. 1, 2017
Universidad del Rosario
Leticia Domingues Bertuzzi
Universidade Federal de Ciências da Saúde de Porto Alegre, Brasil
Patrícia Santos da Silva
Universidade Federal de Ciências da Saúde de Porto Alegre, Brasil
Élida Fluck Pereira Neto
Universidade Federal de Ciências da Saúde de Porto Alegre, Brasil
Rafael Rafael Nicolaidis
Universidade Federal de Ciências da Saúde de Porto Alegre, Brasil
Alcyr Alves de Oliveira Jr
Universidade Federal de Ciências da Saúde de Porto Alegre, Brasil
Received: 03 February 2016
Accepted: 27 June 2016
Additional information
Cómo
citar este artículo: Bertuzzi, L. D.,
Silva, P. P., Fluck, E., Nicolaidis,
R., & Oliveira, A. A. (2017). Animal Model
of Cognitive Decline and Neural Transplantation of Stem Cells as a Prospective
Therapy. Avances en Psicología Latinoamericana, 35(1),
165-175. doi: https://doi.org/10.12804/revistas.urosario.edu.co/apl/a.4665
Abstract: There are few animal models of cognitive decline that could be considered as prominent as the cholinergic lesion model of Alzheimer’s disease. Alzheimer’s disease is a neurodegenerative disorder associated with aging with accentuated progressive cognitive impairment. Its multiple complex sequential and interactive neural pathways may lead to the understanding that a single agent intervention might be insufficient to impact the course of this disease. The current pharmacological treatment is palliative and, although it may slow down the disease’s progression, it still cannot offer cure. The combination of drugs and antioxidant substances has been investigated, nevertheless there are few studies demonstrating optimal and consistent results based on this combination. Thus, the purpose of this paper is to highlight cell-based therapies as prospective treatment of peripheral and central nervous system disorders.
Keywords: Animal models of cognitive decline, Alzheimer’s disease, stem cell based therapy.
Resumen: Existen pocos modelos animales de degeneración cognitiva que podrían ser considerados tan importantes como el modelo de lesión colinérgica de la enfermedad de Alzheimer. La enfermedad de Alzheimer es una patología neurodegenerativa asociada con el envejecimiento y con degeneración cognitiva acentuada y progresiva. Sus complejas y múltiples vías neurales secuenciales pueden conducir a la comprensión de que las intervenciones de agente único pueden ser insuficiente para impactar el curso de esta enfermedad. El tratamiento farmacológico actual es paliativo y, aunque pueda retardar el progreso de la enfermedad, no es capaz de ofrecer una cura. La combinación de drogas y sustancias antioxidantes han sido investigadas, sin embargo, existen pocos estudios que demuestran resultados consistentes basados en esta presunción. Así, el objetivo de este artículo fue destacar las terapias basadas en la reposición celular como potencial tratamiento para enfermedades del sistema nervioso periférico y central.
Palabras clave: modelos animales de degeneración cognitiva, enfermedad de Alzheimer, terapia de células troncales.
Resumo: Existem poucos modelos animais de declínio cognitivo que poderiam ser considerados tão importantes como o modelo de lesão colinérgica da doença de Alzheimer. A doença de Alzheimer é uma patologia neurodegenerativa associada com o envelhecimento e com acentuada e progressiva disfunção cognitiva. Suas complexas e múltiplas vias neurais sequenciais e pode conduzir à compreensão de que as intervenções de agente único pode ser insuficiente para impactar o curso desta doença. O tratamento farmacológico atual é paliativo e, embora possa retardar a progressão da doença, não é capaz de oferecer a cura. A combinação de drogas e substâncias antioxidantes tem sido investigada, no entanto, existem poucos estudos que demonstram resultados consistentes baseados nesta presunção. Assim, o objetivo deste artigo foi destacar as terapias baseadas na reposição celular como potencial tratamento para doenças do sistema nervoso periférico e central.
Palavras-chave: modelos animais de declínio cognitivo, doença de Alzheimer, terapia de células tronco.
In acute cases of neurological damages such as stroke or spinal cord injury, different types of neurons and glial cells die within restricted areas of the central nervous system (CNS) area over a short period of time (Lindvall & Kokaia, 2010). In chronicle situations, there is either selective loss of a specific cell population such as dopaminergic neurons in Parkinson’s disease (PD) or a widespread degeneration of many types of neurons over a period of several years such as in Alzheimer’s disease (AD) (Lindvall & Kokaia, 2010).
Alzheimer’s disease is the most prevalent neurodegenerative disorder associated with aging (Niedowicz, Nelson, & Murphy, 2011) and its incidence is expected to grow intensively such that by 2050 more than 100 million people worldwide will have developed this dementia (Prince, Bryce, & Ferri, 2011). AD is characterized by the progressive loss of cognitive functions, specially, memory. However, besides compromising the memory, AD is also responsible for deficits in attention, orientation, organization, executive functions such as the capacity of planning and judging (Weintraub et al., 2009) leading to the inability of living independently.
Pathological hallmarks of AD are dystrophic neuritis associated with plaques and neurofibrillary tangles within nerve cell bodies in the basal forebrain and neocortical cholinergic pathways (Wszolek, 2008). The widespread loss of neurons and synaptic connectivity in AD seem to be caused by the accumulation of toxic species of the β-Amyloid (Aβ) peptide (Chen & Blurton-Jones, 2012).
It has been reported that the toxic deposit of Aβ causes membrane blebbing and cell shrinkage followed by DNA damage, generation of nuclear apoptotic bodies, DNA fragmentation, and other classic hallmarks of apoptosis (Paradis, Douillard, Koutroumanis, Goodyer, & LeBlanc, 1996). Preventing the apoptosis induced by Aβ and reconstructing the cholinergic circuitry could be an optimal treatment for AD. However, since the relationship among pathological features remains elusive, pharmacological therapies keeps the focus on the function of acetylcholine through cholinesterase inhibitors (ChE-I) while stem cells (SC) researchers look for a cell-based therapy (Oliveira & Hodges, 2005).
Pharmacotherapy
Multiple complex sequential and interactive pathways are involved in the AD’s neurobiology, and interventions with a single agent could be insufficient to impact the disease’s course (Cummings, Gould, & Zhong, 2012). Pharmacological treatment for AD is palliative, having the property of slowing down the speed of the disease’s progression. Several studies have shown that the combination treatment of acetylcholinesterase inhibitor (AChE-I) and memantine prolongs the time an AD’s patient is able to live more independently (Pfeil, Kressig, Thomas, & Szucs, 2012). However, according to Pfeil et al. (2012) these benefits can only be seen after a year of this combination therapy.
Cholinesterase inhibitors increase cholinergic synaptic transmission by holding back acetylcholinesterase in the synaptic cleft, thereby decreasing the hydrolysis of acetylcholine released from the presynaptic neurons (Chu, 2012). Despite the extensive discussion on the effectiveness of AChE-I donepezil, galantamine and rivastigmine, these are the drugs currently approved by the FDA for treating AD’s patients in moderate and mild stages (Mini Mental State Examination scores between 10 and 24), showing a delay in the cognitive impairment and some of the behavioral problems (Sadowsky & Galvin, 2012).
In general, AChE-I drugs are well tolerated; nevertheless, dose-related adverse effects may occur. It is not possible yet to identify which AD patient will respond best to each one of these drugs, nor predict who will develop more intense adverse reactions such as vomiting, nausea and diarrhea, the most prevalent ones (Sadowsky & Galvin, 2012). Meanwhile, studies of AD’s patients treated by rivastigmine, donepezil and galantamine for up to 4 years demonstrated fewer declines in cognitive functions than expected (Chu, 2012).
Some studies have investigated the role of antioxidant treatment in AD. Epicatechin, a polyphenol flavan-3-ol that can be found in food and beverages such as chocolate in natura, red wine, some fruit and teas, especially the traditional green tea Camellia sinensis, can exert neuroprotective properties (Aktas, Waiczies, & Zipp, 2007). Fruit and vegetables rich in flavonoid played an important role in the prevention of motor and cognitive declines related to aging (Bickford, Shukitt-Hale, & Joseph, 1999). In an in vitro experiment, epichatechin’s antioxidant property could be responsible for the prevention of cells deaths by reducing the oxidative stress (Schroeter, Williams, Matin, Iversen, & Rice-Evans, 2000).
Limited data are available considering antioxidant treatments in dementia. According to Weinmann, Roll, Schwarzbach, Vauth and Willich (2010), the extract of Ginkgo Biloba, an herb considered to have potent antioxidant property due to the presence of flavonoids and terpenoids, is not truly recommended for AD’s treatment. Even though in the subgroup of patients with AD there was a significant advantage of Ginkgo Biloba compared to placebo in the conducted study, there was no relevant benefit of the herb in the treatment of neuropsychiatric symptoms. Therefore, additional studies comparing the relative effectiveness of Gingko Biloba and AChE-I should be conducted (Weinmann et al., 2010).
Huperzine-A (Hup-A), an alkaloid isolated from Huperzia serrata, an herb used in the traditional Chinese medicine for treating fevers, swellings, hypertension and memory impairment is considered to be a potent AChE–I proven to effectively improve the learning, memory performance and behavioral dysfunction in Chinese with AD (Gao, Zheng, Yang, Tang, & Zhang, 2009; Wang, Yan, & Tang, 2006). In a study with rats, an isolation of brain mitochondria demonstrated the protective effects of Hup-A on mitochondrial function and on mitochondria’s extended noncholinergic functions as well, providing a significant potential for the design of anti-AD drugs (Gao et al., 2009).
Animal Models for Alzheimer’s Disease
Animal models have been very important to understand the complex pathogenic mechanism underlying neurodegenerative diseases. The use of neurotoxins characterizes an alternative to reproduce an AD model. The cytotoxin infusion of agents capable of destroying cell populations or brain structures is a common method for this (see table 1). Even though the neurotoxins’ actions destroy neuronal cells by a massive postsynaptic depolarization (Barker & Dunnett, 1999), they have their own specificities and limitations.
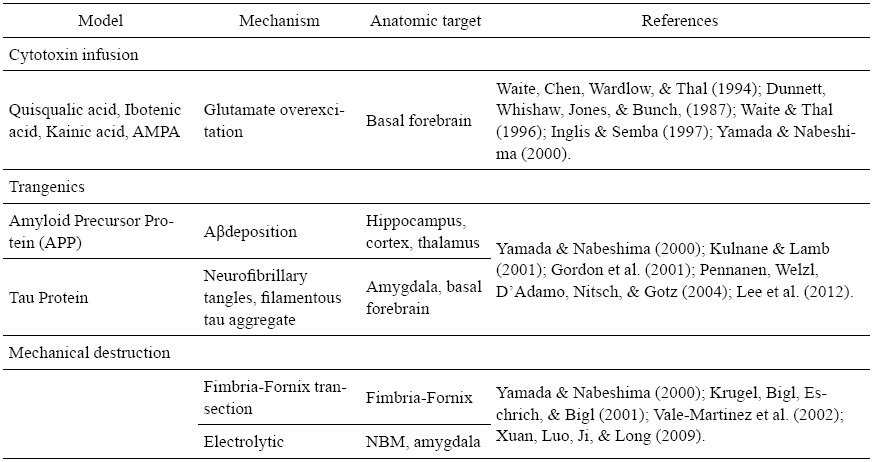
Transgenic models have also been developed, providing considerable information about the AD’s neuropathology. According to Granic, Masman, Luiten and Eisel (2010) transgenic models of AD rely on inherited forms of the disease and present indistinguishable clinical and histopathological features. It is important to highlight that, even though there is a great quantity of transgenic AD models, there is no perfect model that incorporates all aspects of AD and some effective results in the animals fail on clinical trials.
Neuropathway mechanical lesions in the electrolytic model by direct target destruction or fimbria-fornix’ transaction by pathway axotomy can also be an option; nevertheless there are chances of unspecific results (Oliveira & Hodges, 2005).
In addition to the models presented, others can be pointed such as aging animals characterized by an overall age related deficits of neurotransmitter systems. This senescence model favors the natural decline of neurotransmitter systems, however it is not a widespread model due to its long time period keeping and maintaining the animals as well as the animals’ inconsistence cognitive decline (Oliveira & Hodges, 2005).
Stem Cells Therapy for Neurodegenerative Diseases
In the 80’s, researchers believed that neurodegenerative diseases such as AD and PD could benefit from SCs once there would be an increase of the cells in the brain (Gash, Collier, & Sladeck, 1985). Almost 30 years later, stem cell-based therapies have aimed to replace missing or defective cells in many disorders, but a lot is to be known about the mechanism of the SCs inside the organism. Shortly, SCs are classically defined as cells that have the ability to renew themselves continuously and possess totipotent, pluripotent or multipotent ability to differentiate into various cell types (Wislet-Gendebien, Laudet, Neirinckx, & Rogister, 2011). Studies indicate that different lineages of SCs promote a series of changes throughout the recipient’s CNS not restricted only to the specific area (Giusto, Donegá, Cossetti, & Pluchino, 2013; Bonnamain, Neveu, & Naveilhan, 2012).
According to Dunnett, Low, Iversen, Stenevi and Björkland (1982), implanted animals with cholinergically rich cells were able to perform a requested task as the control animals; however, the time required for its execution was much higher than that expended by the control animals. A cholinergic reinnervation is known to be needed; nevertheless in the study it was insufficient to a successful functional recovery (Dunnet et al., 1982). Some years later, ectopic implantation of forebrain ventral cells (cholinergically rich) attenuated learning deficits in rats with cholinergic lesions in a study proposed by Barker and Dunnet (1999).
Modo et al. (2002) reported that the privileged immune characteristics of CNS can be attributed to numerous factors such as the low immunogeneticity of neuronal tissues, the protection of the blood-brain barrier and the absence of lymphatic drainage in the brain. However, CNS immune’s protection is only partial once there are differences in species between the donor and the host and rejection occurs (Modo et al. 2002). Thus, the treatment with immunosuppressive drugs is necessary in cases of allogeneic transplants.
MSCs may be defined as progenitor cells capable of giving rise to a number of unique, differentiated cells from all three germ layers (Caplan, 1991). First described as fibroblast precursors from bone marrow (BM) by Friedenstein, Chailakhjan and Lalykina (1970), MSCs may be also referred to as a fibroblast colony-forming units or marrow stromal cells (Phinney, 2002). Even though these SCs have more limited ability of differentiation than the embryonic SCs, MSCs have important advantages such as the facility for isolation, the capacity to propagate in culture, besides the non-immunogenic characteristics (Bydlowski, Debes, Maselli, & Janz, 2009).
Cova et al. (2010) found that human MSCs promoted an effective neuronal survival and supported the development of immature cells in a PD’s animal model. According to them, neuronal proliferation and maturation were found after the implants in animals that had the striatum injured by 6-hydroxydopamine (6-OHDA). The same was not found in sham animals implanted with the same cell line. The results suggest the existence of a mutual influence between the grafted cells and endogenous neural precursors, becoming important to determine the effect on survival neuronal cells in other brain regions.
Naive human MSCs consistently synthesize a variety of trophic factors that play an important role on the improvement observed in implanted animals (Cova et al., 2010). However, despite the promising outcomes, important issues such as tissue’s viability, logistic and ethical concerns surrounding the use of human material, cell’s purity and viability, deleterious side effects, to site some, need to be better evaluated and understood until this practice becomes commonly used (Barker, Barrett, Mason, & Björklund, 2013).
Bone marrow mesenchymal SCs (BM-MSC) show a great potential of proliferating and differentiating into several types of cells, and present two major advantages over other cell sources: accessibility and immunological features (Oliveira, Haeser, & Pranke, 2008). Besides this, the use of BM-MSC’s for grafts circumvents the ethical concerns attached to the use of embryonic cells. It also brings technical and practical solutions, such as simpler cell collection since it has been practiced in BM transplants.
Recent studies show that BM-MSCs are satisfactory used in intracerebral procedures for neurodegenerative diseases in animal models. According to Lee, Jin and Bae (2009), BM-MSCs promoted microglial activation and reduced significantly the Aβ deposit in a mouse model. Hence, BM-MSCs can increase the number of activated microglia, which supports the hypothesis that BM-MSCs might be an effective therapeutic vehicle to reduce Aβ deposits in AD patients (Lee et al., 2009).
Adipose-derived MSCs (AMSC) isolated from adipose tissue can be differentiated into neuronal SCs, and so improve the feasibility for neurodegenerative diseases’ treatment. Besides this, AMSCs secret anti-apoptotic growth factors, reproducing cytoprotective effects in animal models ( Im et al., 2013). Studies have investigated the neuroprotective effects of the AMSC in animal models of intracerebral hemorrhage and found out evidences of a reduction in the acute cerebral inflammation and in the chronic brain’s degeneration after implantation, which corroborated to improve functional recovery (Kim et al., 2007).
Therapeutic potential of intracerebral injection of human AMSCs in neurodegenerative diseases was previously reported in Huntington’s disease, PD and ischemia mouse models, but not in AD (Kim et al., 2012). In a carefully designed study on therapeutic effects of intravenous human adipose-derived SCs in an AD model, Kim et al. (2012) observed that intravenous or intracerebral human AMSC transplantation rescued memory impairments, besides reducing the number of Aβ plaques in the mice’s brain. There was no notification sign of tumorigenisis in this study; nevertheless Gimble, Bunnell, Chiu and Guilak (2011) identified the formation of sarcomas in vivo after the implantation of AMSCs in immunodeficient mice.
Dental and periodontal tissues constitute a recent source of neural SCs, presenting a substantial advantage among other SCs: their neural crest phenotype (Ibarretxe et al., 2012). Dental pulp SCs (DPSC) and human exfoliated teeth SCs (ETSC) have been shown to differentiate into the three germ layers, also displaying a potency that was widely thought to be exclusive from embryonic SCs (Janebodin et al., 2011; Miura et al., 2003). When compared to BM-MSCs, ETSCs present more active and developed metabolic cells (Karaoz, Sağlam, Aksoy, Kaymaz, & Duruksu, 2011).
These characteristics were corroborated in a study on the differentiation process of human exfoliated deciduous teeth SCs into neurons that performed a dopaminergic function (Wang et al., 2010). As a result, it is estimated that the dental pulp SCs become an alternative source of human MSCs, once they offer promising differentiation properties, high proliferative capacity, and don’t raise ethical concerns as embryonic SCs (Ibarretxe et al., 2012).
Few studies have identified kidney-derived SCs (KSCs), especially considering neurodegenerative diseases. Studies about the isolation of kidney-derived cells showed consistent characteristics of renal SCs, without evidences of cell senescence or transformation into malignant tumors (Gupta et al., 2006). According to Ranghini et al. (2013), studies have shown that some kidney stem/progenitor cells (KSPCs) can differentiate into a variety of renal cell types (Lee et al., 2010).
However, in most cases, differentiation was defined solely on the basis of marker analysis, remaining unknown whether these stem/progenitor cell-derived renal cell types display any functionality. Nevertheless, another study suggested that human KSPCs cells improve renal function by secreting paracrine factors that promote tubular repair (Ranghini et al., 2013; Sallustio et al., 2013).
Conclusions
The development of models for behavior and cognitive decline in neurodegenerative diseases is a constant challenge for comparative psychology researchers. Associate the search for a comprehensive model of a multifactorial condition that may cause cognitive impairment is a methodological hotspot. Although it is difficult sometimes to consider a model of disease as a useful tool for knowledge achievement, models for non-pathological questions might be even more difficult.
The existing animal models for aging pathologies have been used in the development of various treatments and helped to bring discovery for several advances in the health sciences. The models of cognitive impoverishment such as the lesion animal model of Alzheimer’s disease has open fields for the study of memory and attention and is an important tool for the search of its treatment.
A substantial segment of the aging population constitutes the primary risk factor for AD and other neurodegenerative disorders. Intensive efforts have been dedicated to improve old people’s life style habits such as exercise and dietary supplements in combination with symptomatic treatment drugs to improve age-related cognitive decline and to attenuate motor and neurological dysfunction in neurodegenerative diseases (Mandel & Youdim, 2012). However, the adult brain has limited regenerative capacity and no effective treatment options are available (Andresa, Meyerb, Ducraya, & Widmera, 2008).
Stem cells-derived therapy constitute promising prospects for the treatment of CNS disorders, once it is known that SCs have a great potential to develop into many types and subtypes of cells, besides being able of in vitro long-term survival and self-renewal (Ibarretxe et al., 2012). Implants from SCs demonstrated an improvement in the performance of rats in a model for AD in behavioral tests (Bertuzzi et al., n/d). The existence of these behavioral improvement is not exclusively associated to implants survival. Rehabilitation of tissues may occur due to innate mechanisms of the SCs to migrate to the lesion site, in particular, inflammations, apoptosis or hypoxia, releasing trophic factors that accelerate the endogen repair (Joyce et al., 2010).
The migration of the SCs to injured tissues in animal models is associated to a mechanism not yet clearly understood and it is likely involved in the production of paracrine, anti-inflammatory and anti-apoptotic factors (Roux, Leotot, Chevallier, Bierling, & Rouard, 2011). Paracrine effect is defined as a form of communication between two different cells. One releases chemical mediators to its immediate environment, resulting in a change in the behavior of a cell in its adjacent environment (Anthony & Shiels, 2013).
Several experiments have demonstrated that SCs’ grafts ameliorate a variety of CNS conditions. However, crucial issues such as functional connectivity, in vivo differentiation and survival, potential for migration to areas of lesion, host’s immune reactions or mechanisms regulating these functions, are still to be addressed (Oliveira et al., 2008). Existing clinical data suggest that transplantation is technically feasible and can be carried out safely. Nevertheless, data on functional outcome and long-term efficiency remains preliminary (Andresa et al., 2008). A greater understanding of the neurobiological and molecular factors that determine the progress of the brain disorder will probably lead to safer treatments (Oliveira et al., 2008).
Stem cells based therapies represent an emerging therapeutic approach by which incurable diseases may be treated (Marchetti, Krohne, Friedlander, & Friedlander, 2010). This search is deeply influenced by the development of new models and the comprehension of the relationship between behavior, cognition and its biological somatic correspondent.
In neural transplantation, although more should be known to clarify all the mechanisms of cellular paracrine effects in the host’s tissues, there is a good perspective that cell therapies being played in the right models might be used to treat neurodegenerative diseases.
References
Aktas, O., Waiczies, S., & Zipp, F. (2007). Neurodegeneration in autoimmune demyelination: recent mechanistic insights reveal novel therapeutic targets. Journal of Neuroimmunology, 184(1-2), 17-26.
Andresa, R. H., Meyerb, M., Ducraya, A. D., & Widmera, H. R. (2008). Restorative neuroscience: concepts and perspectives. Swiss Medical Weekly, 138(11-12), 155-172.
Anthony, D. F. & Shiels, P. G. (2013). Exploiting paracrine mechanisms of tissue regeneration to repair damaged organs. Transplantation Research, 2(1), 10.
Barker, R. A., Barrett, J., Mason, S. L., & Björklund, A. (2013). Fetal dopaminergic transplantation trials and the future of neural grafting in Parkinson’s disease. The Lancet Neurology, 12(1), 84-91.
Barker, R. A. & Dunnet, S. B. (1999). Neural repair, transplantation and rehabilitation. Hove: Psychology Press.
Bertuzzi, L., Rosa, A. R., Ourique, A., Wendt, L., Nicolaidis, R., Torquato, K., Stapenhorst, A., Silva, P. S., Fluck, E., Chagas Telles, P., Pranke, P., & Oliveira, A. A. (n/d). Stem cell’s behavioral effects in rats in a model of Alzheimer’s disease. Advances in Stem Cells. doi: 10.5171/2014.805238
Bickford, P.C., Shukitt-Hale, B., & Joseph, J. (1999). Effects of aging on cerebellar noradrenergic function and motor learning: nutritional interventions. Mechanisms of Ageing and Development, 111(2-3), 141-154.
Bonnamain, V., Neveu, I., & Naveilhan, P. (2012). Neural stem/progenitor cells as promising candidates for regenerative therapy of the central nervous system. Frontiers in Cellular Neuroscience, 6(17). doi: 10.3389/fncel.2012.00017
Bydlowski, S. P., Debes, A. A., Maselli, L. M. F., & Janz, F. L. (2009). Características biológicas das células-tronco mesenquimais. Revista Brasileira de Hematologia e Hemoterapia, 31 (supl.1), 25-35.
Caplan, A. I. (1991). Mesenchymal stem cells. Journal of Orthopaedic Research, 9, 641-650.
Chen, W. W. & Blurton-Jones, M. (2012). Concise review: Can stem cells be used to treat or model Alzheimer’s disease? Stem Cells, 30, 2 612-2 618.
Chu, L.W. (2012). Alzheimer’s disease: Early diagnosis and treatment. Hong Kong Medical Journal, 18, 228-237.
Cova, L., Armentero, M.-T., Zennaro, E., Calzarossa, C., Bossolasco, P., Busca, G. Deliliers, G. L., Polli, E., Nappi, G., Silani, V., & Blandini, F. (2010). Multiple neurogenic and neurorescue effects of human mesenchymal stem cell after transplantation in an experimental model of Parkinson’s disease. Brain Research, 1 311, 12-27.
Cummings, J., Gould, H., & Zhong, K. (2012). Advances in designs for Alzheimer’s disease clinical trials. American Journal of Neurodegenerative Disease, 1(3), 205-216.
Dunnet, S. B., Low, W. C., Iversen, S. D., Stenevi, U., & Björkland, A. (1982). Septal transplants restore maze learning in rats with fornix-fimbria lesions. Brain Research, 251, 335-348.
Dunnett, S. B., Whishaw, I. Q., Jones, G. H., & Bunch, S. T. (1987). Behavioural, biochemical and histochemical effects of different neurotoxic amino acids injected into nucleus basalis magnocellularis of rats. Neuroscience, 20(2), 653-669.
Friedenstein, A. J., Chailakhjan, R. K., & Lalykina, K. S. (1970). The development of fibroblast colonies in monolayer cultures of guinea-pig bone marrow and spleen cells. Cell and Tissue Kinetics, 3, 393-403.
Gao, X., Zheng, C. Y., Yang, L., Tang, X. C., & Zhang, H. Y. (2009). Huperzine A protects isolated rat brain mitochondria against beta-Amyloid peptide. Free Radical Biology and Medicine, 46, 1 454-1 462.
Gash, D. M., Collier, T. J., & Sladek, J. R. (1985). Neural transplantation: A review of recent developments and potential applications to the aged brain. Neurobiology of Aging, 6, 131-150.
Gimble, J., Bunnell, B., Chiu, E., & Guilak, F. (2011). Adipose-derived stromal vascular fraction cells and stem cells: Let’s not get lost in translation. Stem Cells, 29, 749-754.
Giusto, E., Donegà, M., Cossetti, C., & Pluchino, S. (2013). Neuro-immune interactions of neural stem cell transplants: from animals’ disease models to human trials. Experimental Neurology, 13, 92-97.
Gordon, M. N., King, D. L., Diamond, D. M., Jantzen, P. T., Boyett, K. V., & Hope, C. E. (2001). Correlation between cognitive deficits and A(beta) deposits in transgenic APP+PS1 mice. Neurobiology of Aging , 22(3), 377-385.
Granic, I., Masman, M. F., Luiten, P. G. M., & Eisel, U. L. M. (2010). Braak staging in mouse models of Alzheimer’s disease. American Journal of Pathology, 177(4), 1 603-1 605.
Gupta, S., Verfaillie, C., Chmielewski, D., Kren, S., Eidman, K., Connaire, J., Yves, H., Lund, T., Blackstad, M., Jiang, Y., Luttun, A., & Rosenberg, M. E. (2006). Isolation and characterization of kidney-derived stem cells. Journal of the American Society of Nephrology, 17, 3 028-3 040.
Ibarretxe, G., Crende, O., Aurrekoetxea, M., Garcıa-Murga, V., Etxaniz, J., & Unda, F. (2012). Neural crest stem cells from dental tissues: A new hope for dental and neural regeneration. Stem Cells International, 12.
Im, W ., Ban, J ., Lim, J ., Lee, M ., Lee, S. T ., Chu, K ., & Kim, M. (2013). Extracts of adipose derived stem cells slows progression in the R6/2 model of Huntington’s disease. PLoS One, 8(4), e59438.
Inglis, W. L. & Semba, K. (1997). Discriminable excitotoxic effects of ibotenic acid AMPA, NMDA and quinolinic acid in the rat laterodorsal tegmental nucleus. Brain Research, 755(1), 17-27.
Janebodin, K., Horst, O. V., Ieronimakis, N., Balasundaram, G., Reesukumal, K., Pratumvinit, B., & Reyes, M. (2011). Isolation and characterization of neural crest-derived stem cells from dental pulp of neonatal mice. PLoS One, 6(11).
Joyce, N ., Annett, G ., Wirthlin, L ., Olson, S ., Bauer, G ., & Nolta, J. A . (2010). Mesenchymal stem cells for the treatment of neurodegenerative disease. Regenerative Medicine, 5(6), 933-946.
Karaöz, E. D. P., Sağlam, O., Aksoy, A., Kaymaz, F., & Duruksu, G. (2011). Human dental pulp stem cells demonstrate better neural and epithelial stem cell properties than bone marrow-derived mesenchymal stem cells. Histochemistry and Cell Biology. Retrieve from http://www.springerlink.com/content/0948-6143
Kim, J . M., Lee, S . T., Chu, K ., Jung, K. H., Song, E . C., Kim, S. J., Sinn, D. I., Kim, J. H., Park, D. K., Kang, K. M., Hyung, H., Park, N., Won, H. K., Kim, C. H., Kim, K. H. M., Lee, K. S., & Roh, J. K . (2007). Systemic transplantation of human adipose stem cells attenuated cerebral inflammation and degeneration in a hemorrhagic stroke model. Brain Research, 5(1 183), 43-50 .
Kim, S., Chang, K. A., Kim, J., Park, H. G., Ra, J. C., Kim, H. S., & Suh, Y. H. (2012). The preventive and therapeutic effects of intravenous human adipose-derived stem cells in Alzheimer’s disease mice. PLoS One, 7(9).
Krugel, U., Bigl, V., Eschrich, K., & Bigl, M. (2001). Deafferentation of the septo-hippocampal pathway in rats as a model of the metabolic events in Alzheimer’s disease. The International Journal of Developmental Neuroscience, 3, 263-277.
Kulnane, L. S. & Lamb, B. T. (2001). Neuropathological characterization of mutant amyloid precursor protein yeast artificial chromosome transgenic mice. Neurobiology of Disease, 8(6), 982-992.
Lee, J., Jin, H., & Bae, J. (2009). Bone marrow-derived mesenchymal stem cells reduce brain amyloid-B deposition and accelerate the activation of microglia in an acutely induced Alzheimer’s disease mouse model. Neuroscience Letters, 450, 136-141.
Lee, P. T., Lin, H. H., Jiang, S. T., Lu, P. J., Chou, K. J., Fang, H.-C., Chiou, Y.-Y., & Tang, M.-J. (2010). Mouse kidney progenitor cells accelerate renal regeneration and prolong survival after ischemic injury. Stem Cells, 28, 573-584.
Lee, H. J., Lee, J. K., Lee, H., Carter, J. E., Chang, J. W., Yang, Y. S., Suh, J.-G., Lee, B.-H., Jin, H. K., & Bae, J.-S. (2012). Human umbilical cord blood-derived mesenchymal stem cells improve neuropathology and cognitive impairment in Alzheimer’s disease mouse model through modulation of neuroinflammation. Neurobiology of Aging, 33, 588-602.
Lindvall, O. & Kokaia, Z. (2010). Stem cells in human neurodegenerative disorders — time for clinical translation? Journal of Clinical Investigation, 120(1), 29-40.
Mandel, S. A . & Youdim, M. B. (2012). In the rush for green gold: Can green tea delay age-progressive brain neurodegeneration? Recent Patents on CNS Drug Discovery, 7(3), 205-217.
Marchetti, V., Krohne, T. U., Friedlander D. F., & Friedlander, M. (2010). Stemming vision loss with stem cells. Journal of Clinical Investigation, 120(9), 3 012-3 021.
Miura, M., Gronthos, S., Zhao, M., Lu, B., Fisher, L. W., Robey, P. G., & Shi, S. (2003). SHED: stem cells from human exfoliated deciduous teeth. Proceedings of the National Academy of Sciences of the United States of America, 100(10), 5 807-5 812.
Modo, M., Rezaie, P., Heuschling, P., Patel, S., Male, D. K. & Hodges, H. (2002). Transplantation of neural stem cells in a rat model of stroke: assessment of short-term graft survival and acute host immunological response. Brain Research, 958(1), 70-82.
Niedowicz, D. M., Nelson, P. T., & Murphy, P. M. (2011). Alzheimer’s disease: Pathological mechanisms and recent insights. Current Neuropharmacology, 9, 674-684.
Oliveira, A. A., Haeser, A., & Pranke, P. (2008). Stem cell grafts as therapeutic tools for central nervous system disorders. Psychology & Neuroscience, 1(1), 47-54.
Oliveira, A. A. & Hodges, H. (2005). Alzheimer’s disease and neural transplantation as prospective cell therapy. Current Alzheimer Research, 2(1), 79-95.
Paradis, E., Douillard, H., Koutroumanis, M., Goodyer, C., & LeBlanc, A. (1996). Amyloid B Peptide of Alzheimer’s Disease Downregulates Bcl-2 and Upregulates Bax Expression in Human Neurons. Journal of Neuroscience, 16(23), 7 533-7 559.
Pennanen, L., Welzl, H., D’Adamo, P., Nitsch, R. M., & Gotz, J. (2004). Accelerated extinction of conditioned taste aversion in P301L tau transgenic mice. Neurobiology of Disease, 15(3), 500-509.
Pfeil, A. M., Kressig, R. W., Thomas, D., & Szucs, T. D. (2012). Alzheimer’s dementia: Budget impact and cost-utility analysis of a combination treatment with a cholinesterase inhibitor and memantine in Switzerland. Swiss Medical Weekly, 142, w13676.
Phinney, D.G. (2002). Building a consensus regarding the nature and origin of mesenchymal stem cells. Journal of Cellular Biochemistry, 38, 7-12.
Prince, M., Bryce, R., & Ferri, C. (2011). World Alzheimer Report 2011: The benefits of early diagnosis and intervention. Retrieve from http://www.alz.co.uk .
Ranghini, E., Mora, C. F., Edgar, D., Kenny, S. E., Murray, P., & Wilm, B. (2013). Stem cells derived from neonatal mouse kidney generate functional proximal tubule-like cells and integrate into developing nephrons in vitro. PLoS One, 8(5), e62953.
Roux, S ., Leotot, J ., Chevallier, N ., Bierling, P ., & Rouard, H . (2011). Mesenchymal stromal cells: Biological properties and clinical prospects. Transfusion Clinique et Biologique, 18(1), 1-12.
Sadowsky, C. H. & Galvin, J. E. (2012). Guidelines for the management of cognitive and behavioral problems in dementia. American Board of Family Medicine, 25(3), 3 350-3 366.
Sallustio, F., Costantino, V., Cox, S. N., Loverre, A., Divella, C., Rizzi, M., & Schena, F. P. (2013). Human renal stem/progenitor cells repair tubular epithelial cell injury through TLR2-driven inhibin-A and microvesicle-shuttled decorin. Kidney International, 83(3), 92-103.
Schroeter, H., Williams, R. J., Matin, R., Iversen, L., & Rice-Evans, C. A. (2000). Phenolic antioxidants attenuate neuronal cell death following uptake of oxidized low-density lipoprotein. Free Radical Biology and Medicine, 29(12), 1 222-1 233.
Vale-Martinez, A., Guillazo-Blanch, G., Marti-Nicolovius, M., Nadal, R., Arevalo-Garcia, R., & Morgado-Bernal, I. (2002). Electrolytic and ibotenic acid lesions of the nucleus basalis magnocellularis interrupt long-term retention, but not acquisition of two-way active avoidance, in rats. Experimental Brain Research, 142(1), 52-66.
Waite, J. J. & Thal, L. J. (1996). Lesions of the cholinergic nuclei in the rat basal forebrain: excitotoxins vs. an immunotoxin. Life Sciences, 58(22), 1 947-1 953.
Waite, J. J., Chen, A. D., Wardlow, M. L., & Thal, L. J. (1994). Behavioral and biochemical consequences of combined lesions of the medial septum/diagonal band and nucleus basalis in the rat when ibotenic acid, quisqualic acid, and AMPA are used. Exp Neurol, 130(2), 214-229.
Wang, J., Wang, X., Sun, Z., Wang, X., Yang, H., Shi, S., & Wang, S. (2010). Stem cells from human-exfoliated deciduous teeth can differentiate into dopaminergic neuron-like cells. Stem Cells and Development, 19(9), 1 375-1 383.
Wang, R., Yan, H., & Tang, X. C. (2006). Progress in studies of Huperzine-A, a natural cholinesterase inhibitor from Chinese herbal medicine. Acta Pharmacol Sin, 27, 11-26.
Weinmann, S., Roll S., Schwarzbach, C., Vauth, C., & Willich, S. N. (2010). Effects of Ginkgo biloba in dementia: systematic review and meta-analysis. BMC Geriatrics, 10(14), 12.
Weintraub, S., Salmon, D., Mercaldo, N., Ferris, S., Graff-Radford, N. R., Chui, H., Cummings, J., DeCarli, C., Foster, N., Galasko, D., Peskind, E., Dietrich, W., Beekly, D., Kukull, W. A., & Morris, J. C. (2009). The Alzheimer’s Disease Center’s Uniform Data Set (UDS): The neurpsychological test battery. Alzheimer Disease and Associated Disorders, 23(2), 91-101.
Wislet-Gendebien, S., Laudet, E., Neirinckx, V., & Rogister, B. (2011). Adult bone marrow: Which stem cells for cellular therapy protocols in neurodegenerative disorders? Journal of Biomedicine and Biotechnology, 2012, 10.
Wszolek, C. W. Z. K. (2008). Etiology and pathophysiology of frontotemporal dementia, Parkinson disease and Alzheimer disease: Lessons from genetic studies. Neurodegenerative Disease, 5, 122-125.
Xuan, A. G., Luo, M., Ji, W. D., & Long, D. H. (2009). Effects of engrafted neural stem cells in Alzheimer’s disease rats. Neuroscience Letters, 450, 167-171.
Yamada, K. & Nabeshima, T. (2000). Animal models of Alzheimer’s disease and evaluation of anti-dementia drugs. Pharmacology & Therapeutics, 88, 93-113.




